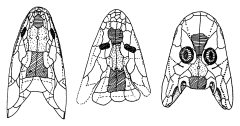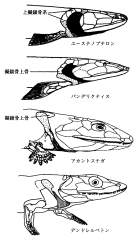 
ユーステノプテロン (上)・パンデリクティス (中上)・アカントステガ (中下)・デンドレルペトン (下) の肩帯から頭部にかけての形態。
魚類から陸上四肢動物への進化の中で,胴体と頭部の間に首ができた。
(Clack, J. A. 手足を持った魚たち――脊椎動物の上陸戦略. 池田比佐子訳. 東京, 講談社, 2000. p.144)
|
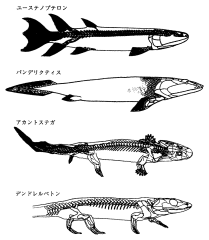
ユーステノプテロン (上)・パンデリクティス (中上)・アカントステガ (中下)・デンドレルペトン (下) の体骨格。パンデルクティスは,まだ体骨格のつくりが完全には明らかにされていない。
(Clack, J. A. 手足を持った魚たち――脊椎動物の上陸戦略. 池田比佐子訳. 東京, 講談社, 2000. p.156)
|